
Следующим очень важным этапом размножения цветковых является опыление- перенос пыльцевых зерен на рыльце пестика (напомнить, что у голосеменных пыльцевые зерна при опылении попадают непосредственно на семязачаток). Обращаем особое внимание на огромное разнообразие у цветковых растение адаптаций, обеспечивающих или облегчающих опыление. Пыльца переносится на рыльце пестика различными способами, но в зависимости от того, с какого цветка эта пыльца берется, выделяют два основных типа: самоопыление и перекрестное опыление.
У перекрестноопыляемых цветковых растений очень разнообразны. Условно их можно разделить на две группы. В первой перенос пыльцы осуществляется животными, главным образом, насекомыми, а также некоторыми позвоночными (большинство из которых также освоили воздушную среду — птицы и летучие мыши). Во второй группе пыльца переносится абиотическими факторами — ветром и реже водой.
У растений, опыляемых животными, обычно формируются различные адаптации, способствующие привлечению опылителей. Обычно они имеют яркоокрашенные крупные цветки. Если цветки мелкие, то они, как правило, оказываются собранными в соцветия, что также зрительно увеличивает их. Однако размеры, окраска и форма цветка позволяют опылителю только увидеть его. Для стимуляции животного исполнить роль опылителя обычно недостаточно, поэтому растения наряду с этим привлекают еще и пищей, которой чаще всего является нектар, выделяемый нектарниками (более подробно о нектарниках рассказано в разделе, посвященном выделительным тканям). Некоторые растения также образуют большое количество пыльцы, которую опылители охотно поедают. Необходимо, чтобы пыльца любым способом попала на опылителя, который переносит ее на другой цветок этого же вида.
Биотическими опылителями являются насекомые, птицы и млекопитающие. Соответственно различают энтомофилию, орнитофилию и зоофилию.
Энтомофилия, или опыление насекомыми, чрезвычайно широко распространено в природе. Роль насекомых в историческом развитии покрытосеменных трудно переоценить, однако и сами насекомые многим обязаны цветковым. В итоге наблюдается большое разнообразие взаимных адаптаций, которые выработали растения и насекомые. Эти адаптации порой бывают настолько узкими, что растение может опыляться лишь определенным видом насекомых.
Чаще всего растение привлекает насекомых пищей — нектаром или пыльцой. При этом нектар выделяется именно в то время суток, когда опылитель активен — днем или ночью. Цветки таких растений обычно крупные и яркие. Однако насекомые зачастую воспринимают окраску цветков совсем иначе, нежели человек, поскольку они воспринимают свет, не доступный нам (например, ультрафиолетовые лучи). Это обстоятельство хорошо известно пчеловодам, которые могут ночью осматривать улей, пользуясь для освещения фонарем, используемым фотографами при проявке фотопленок и печатании снимков. Этот фонарь излучает длинноволновый красный свет, которые пчелы не видят, но видит человек. Если бы пчеловод использовал при этом фонарь с синим светом, то он был бы неминуемо опознан пчелами и искусан. По мнению ботаников, незаметные для человека обозначения на частях цветка служат для насекомого указателем месторасположения нектарников, т. е. цветок представляет собой своеобразную карту, понятную опылителям. Многих насекомых привлекает не только (или не столько) нектар, но и пыльца. Энтомофильные растения обычно вырабатывают пыльцу в большом количестве, кроме того, отдельные пылинки имеют шероховатую поверхность наружной оболочки, что способствует их прилипанию к покровам насекомого. Часто пыльца скатывается в комочки — поллинии (рис. 300), которые и прилипают к телу опылителя (например, у орхидных). Те же пчелы активно собирают пыльцу (рис. 301) и переносят ее в гнездо на своих лапках (обножки). Там они отделяют комочки пыльцы, складывают ее в ячейки и утрамбовывают головой. Когда часть ячейки оказывается частично заполненной, пыльца заливается медом и . В результате брожения углеводов меда образуется молочная кислота, которая обладает консервирующим действием, из-за чего пыльца может долго храниться. Обработанная таким образом пыльца называется пергой. Она очень богата белками, что особенно важно для пчел, поскольку основу их рациона составляют углеводы нектара. Понятно, что растения, опыляемые днем, имеют более яркие цветки, тогда как цветки «ночных» растений обычно окрашены в светлые тона — белые, желтые, светло-красные и т.д., чтобы выглядеть контрастно на общем темном фоне. Интересно, что многие ночные насекомые обладают цветовым зрением и хорошо различают не только яркость, но и оттенки. Цветки привлекают насекомых не только внешним видом, но и запахом. Обычно для этого выделяются эфирные масла, которые представляют собой сложную смесь различных органических соединений (спирты, фенолы, альдегиды, эфиры терпены и т.д.). Все эфирные масла летучи, им свойствен резкий и чаще всего приятный запах.
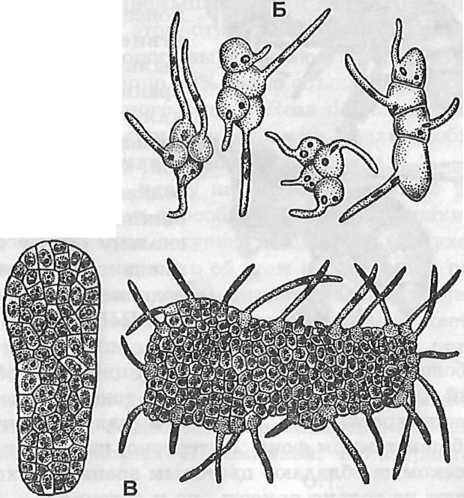
Рис. 300. Пыльца некоторых орхидных, соединенная в тетрады и поллинии, и ее прорастание:
А — венерин башмачок (Cypripedium insigne): Б — каланта вейча (Calanthe veitchii); В — любка двулистная — Platanthera bifolia (по Поддубной-Арнольди)
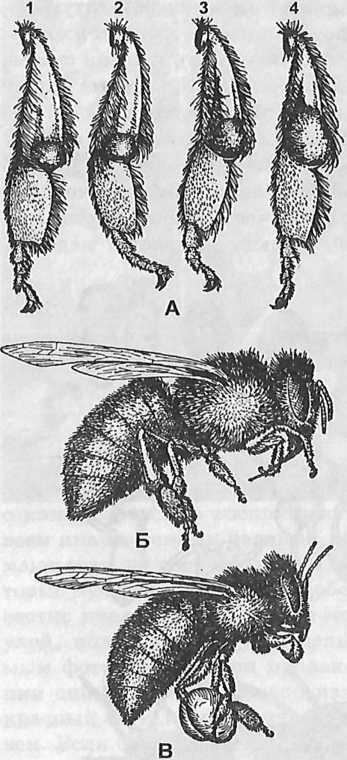
Рис. 301. Сбор пыльцы: А- последовательные фазы (1,2,3,4) наполнения корзиночки пыльцой (обножкой); Б- препровождение пыльцы со щеточек в корзиночки во время полета пчелы; В- пчела во время полета с обножкой в корзиночках задних ног. Средними ногами пыльцевой комочек как бы оформляется.
Среди составных компонентов наиболее распространены монотерпены (С10Н16), причем как алифатические, так и циклические. Примером алифатических (ациклических) является линалоол, который придает цветкам ландыша характерный запах. Близкие по химическому строению к линалоолу гераниол, цитронеллол и нерол обеспечивают запах цветкам розы. Коричный спирт определяет запах гиацинта, кетон пармон — фиалки, смесь из линалоола, индола, бензилацетата и жасмона — жасмина и т.д. Эфирные масла широко используются человеком в парфюмерной промышленности и в медицине. Наиболее известно в этом отношении розовое масло, для производства которого выращиваются обширные плантации роз. Примерно 80% всего объема розового масла получается из одного вида — Rosa damascena, причем для получения одного килограмма масла необходимо переработать 35 млн. розовых лепестков. Установлено, что у насекомых обоняние развито очень хорошо, они способны различать запахи, даже если концентрация летучего вещества в воздухе крайне мала (более подробно об этом рассказано в разделе, посвященном насекомым), поэтому насекомые легко находят цветки. Поскольку отдельные части цветка источают разные запахи, насекомые легко их определяют и более легко ориентируются в цветке. Отдельно следует выделить растения, цветки которых выделяют вещества, напоминающие половые аттрактанты насекомых.

Рис. 3 0 3. Опыление офриса насекомоносного (Ophrys insectifera): 1 — общий вид растения; 2 — цветок; 3, 4 — оса в процессе псевдокопуляции; 5 — голова осы с прикрепившимися к ней поллинариями (по В. Н. Гпадковой)
это те вещества, которые выделяют самки насекомых в период размножения для привлечения самцов. Прилетевшие на запах возбужденные самцы пытаются копулировать с цветком (который вдобавок ко всему еще имеет форму насекомого) и при этом покрываются пыльцой, которую они потом переносят на другой цветок. Такие адаптации выработали некоторые орхидные, опыляемые одиночными перепончатокрылыми (рис. 303).
Таким образом, у растений имеется дальняя сигнализация, стимулирующая прилет опылителя, и ближняя сигнализация, стимулирующая посещение опылителем цветка. Однако у разных насекомых дальняя и ближняя сигнализация воспринимается разными органами чувств. Например, у дневных опылителей прилет стимулируется внешним видом цветка, его размерами, окраской, формой, определяемыми зрительно, тогда как у ночных насекомых фактором дальнего привлечения является запах. Например, цветки душистого табака опыляются ночными бабочками. Поэтому они днем закрыты, а ночью распускаются и усиливают аромат. Запах привлекает издалека не только ночных насекомых, но и многих дневных. В особенности это относится к цветкам, которые источают запах гниющего мяса или экскрементов (например, раффлезии, аронники, стапе- лии). Такие цветки привлекают тучи мух, которые и опыляют цветки. Как мы уже говорили, растения вырабатывают морфологические адаптации, которые обеспечивают попадание пыльцы на опылителя. Обычно строение цветка таково, что, добираясь до желанного нектара, опылитель покрывается пыльцой. Перелетая затем на другие цветки, он трется измазанным телом о рыльце, и пыльцевые зерна прилипают к нему. Нередко цветок при этом имеет очень сложное строение и вынуждает насекомое проделать движения, необходимые для попадания на него пыльцы (рис. 304).
Тесное и взаимовыгодное сотрудничество насекомых и покрытосеменных оказалось чрезвычайно плодотворным. Поэтому в настоящее время цветковые доминируют в царстве растений и имеют самое значительное видовое разнообразие. Это же относится и к насекомым, численность видов которых значительно превышает количество видов всех остальных животных, вместе взятых.
Орнитофилия, т.е. перенос пыльцы птицами-опылителями, распространена менее широко, чем энтомофилия. Чаще всего опыление производят мелкие птицы, такие как колибри (рис. 305) и цветочницы в Америке, нектарницы, белоглазки и медососы в Евразии, гавайские цветочницы. Такие птицы обычно собирают нектар в полете, для чего они зависают в воздухе наподобие бабочек-бражников (размеры колибри могут быть даже меньше крупных бабочек) и исследуют цветки с помощью очень длинного клюва. Если цветок крупный, то птица попросту садится на него. Опылителями могут быть и довольно крупные птицы, например попугаи лори, обитающие в тропической Азии. У этих попугаев очень своеобразный язык — его кончик представляет собой кисточку, с помощью которого птица поглощает нектар. Нектар орнитофильных растений гораздо более жидкий, чем у энтомофильных (содержание сахара всего
около 5%), но его образуется очень много.
Орнитофилия в основном распространена у тропических и экваториальных растений, однако встречается и у растений умеренного климата. Систематическое положение их различно — они относятся как к двудольным (миртовые, бобовые, вересковые и др.), так и к однодольным (банановые, лилейные, бромелиевые и др.).
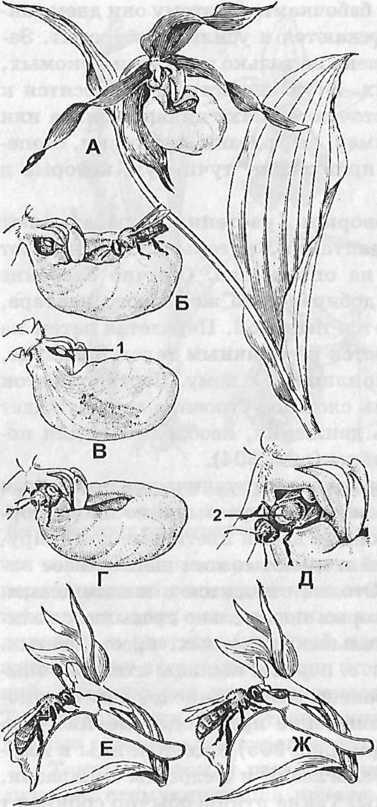
Рис. 304. Опыление орхидейных: башмачок настоящий: А- верхушка побега; Б- насекомое садиться на край губы и соскальзывает внутрь ее; В – насекомое ползет а направлении «окон» (1- комок пыльцы-поллиний); Г – голова насекомого показалась из узкого отверстия расположенного в потную к пыльнику; Д- насекомое с комком липкой пыльцы (2) на спине; пальчатокоренник освистый; Е – насекомое извлекает поллиний; Ж – насекомое вводит поллиний, наклонившийся во время полета в рыльцевую ямку.
Самоопыление встречается у относительно небольшого числа цветковых. В этом случае не происходит обмен генетической информацией между разными особями, поскольку пыльцевые зерна попадают на рыльце пестика либо из пыльника одной из тычинок этого же цветка, либо с другого цветка, расположенного на том же растении. Поскольку все события происходят в пределах одной особи, при самоопылении не происходит обмен генетической информации, а имеют место лишь комбинативные изменения наследственного материала в ходе соответствующих процессов в мейозе (при спорогенезе). Отсутствие новых аллелей приводит к появлению чистых линий гомозиготных популяций в пределах одного вида, неспособных обмениваться генами, подвергшимися мутациям (в том числе полезных), поэтому процессы видообразования в этих популяциях идут самостоятельно.
Постоянное самоопыление возникает при физической невозможности (в силу каких-либо объективных причин) переноса пыльцы с одного растения на другое, при этом обмен растениями пыльцой в обычных условиях не происходит. Такой тип опыления можно рассматривать как адаптацию, поскольку для растительного организма не доступна пыльца с других растений и использование собственных пыльцевых зерен представляется единственной возможностью образовать семена. Случайное самоопыление происходит у многих покрытосеменных в том случае, когда цветки обмениваются между собой пыльцой, но наряду с этим на рыльце возможно попадание пыльцевых зерен, образованных в собственном цветке. В этом случае образуется относительно небольшой процент гомозиготных организмов. Наконец у ряда покрытосеменных в обычных условиях рыльце опыляется чужой пыльцой, но если по каким-либо причинам этого долго не происходит, в последний момент (когда плодолистик еще сохраняет способность воспринимать пыльцевые зерна) на рыльце попадает собственная пыльца, запас которой все это время имеется в цветке. В этом случае самоопыление представляет собой вынужденное событие, но это также представляет собой адаптацию, поскольку совершенно очевидно, что с точки зрения биологической целесообразности лучше произвести самоопыление и сформировать плоды семенами, чем не опылиться вовсе и, соответственно, остаться без семян. Отметим , что самоопылением считается не только перенос с тычинки на рыльце того же самого цветка, но и опыление пыльцой другого цветка, расположенного на том же растении. Причем последний способ самоопыления (он называется гейтеногамией) является единственным возможным у форм с однополыми цветками, поскольку у них тычинки с пыльниками и плодолистики с рыльцами находятся в разных цветках.
Перекрестное опыление, или ксеногамия (греч. хеnos чужой, gamos — брак), представляет собой перенос различными способами пыльцы из пыльника одного растения на рыльце другого. Этот тип опыления у цветковых значительно более распространен, чем самоопыление. В этом случае между разными особями одного вида обязательно происходит обмен аллелями, что приводит к увеличению доли гетерозиготных организмов. Это справедливо можно рассматривать как преимущество по сравнению с самоопылением, поскольку здесь не происходит генетической изоляции отдельных клонов, а возникшие мутации свободно распространяются в пределах популяции.
Для того чтобы произошло перекрестное опыление цветки выработали различные адаптации, которые либо вообще исключают саму возможность самоопыления, либо в какой-то степени ограничивают её вероятность. Следует отметить, что полное отсутствие самоопыления не так уж и полезно для вида, поскольку в этом случае растение оказывается неспособным произвести семена, если по каким-либо причинам не произошло перекрестное опыление(например, отсутствие соответствующих опылителей, низкая плотность произрастания особей данного вида и др.). Для растения наиболее целесообразным является приобретение таких приспособлений, которые до последнего момента будут способствовать перекрестному опылению, но если оно не произойдет, обеспечат самоопыление, и растение при этом сможет образовать семена (пусть даже используя для этого лишь свой генетический материал). Приспособления, ограничивающие самоопыление, очень разнообразны. Рассмотрим некоторые из них.
Двудомность является наиболее надежным способом предотвращения самоопыления. Иными словами, у двудомных растений самоопыление в принципе невозможно, поскольку мужские женские цветки находятся на разных растениях (мужских и женских). Однако не следует считать, что такие растения приобрели идеальную адаптацию. Действительно, у них всегда происходит полноценная комбинация генетического материала и обмен аллелями, но у двудомных растений семена производят лишь женские организмы, так как мужские лишь продуцируют пыльцу. К двудомным растениям относят тополь, иву, осину, коноплю и др.
Своеобразным компромиссом между «достоинствами» и «недостатками» одно- и двудомных растений представляются описанные выше растения, у которых на одной особи развиваются не гермафродитные, а однополые цветки. Однако у них велика вероятность опыления пыльцой с мужских цветков собственного растения. У некоторых растений на одной особи могут развиваться как гермафродитные, так и однополые цветки. Если при этом образуются обоеполые и только мужские (но не образуются женские) цветки, то это называется андромоноэцием (греч. andros — мужчина, monos — один, единый, единственный,oikia — дом, жилище). Противоположное сочетание (обоеполые и только женские цветки) называется гиномоноэцием (греч. gine – женщина, monos – один, единый, единственный, oikia – дом, жилище). Возможны также и другие варианты, например, на одной особи развиваются только двудомные, а на другой только мужские цветки – андродиэций (греч. Andros – мужчина, di – дважды,oikia – дом, жилище). Или, наоборот, на одной особи только обоеполые, а на другой – только женские цветки – гинодиэция (греч. gyne – женщина, di – дважды, oikia – дом, жилище). Гинодиэции в природе распространены более широко, поскольку они позволяют свести к минимуму мужские цветки максимально увеличить численность женских, способных образовывать плоды с семенами. Наконец у ряда растений образуются цветки, имеющие строение обоеполых, но полного развития на одних из них достигают или только тычинки, или только плодолистики.
Дихогамия выражается в неодновременном функциональном развитии тычинок и рыльца в одном цветке. В зависимости от того, что созревает раньше, выделяют протандрию (греч. protos – первый, Andros – мужчина)(раньше созревают пыльники с пыльцой) и протогинию (греч. protos – первый, gyne – женщина)(рыльца созревают раньше пыльников). Протандрия встречается чаще, она имеет место у большого количества семейств цветковых, например у сложноцветковых, зонтичных, гвоздичных, колокольчиковых и др. Гораздо реже встречаются протогиния, более всего она выражена у ветроопыляемых (злаковых, ситиновых, осоковых, подорожниковых и др., но возможна и у розовых,крестоцветных,жимолостных и др.).
Самонесовместимость является наиболее совершенной адаптацией против самоопыления. В этом случае полноценное развитие пыльцевых зерен, попавших на рыльце собственного цветка, становится невозможным. При этом пыльца либо не прорастает вовсе, либо рост пыльцевых трубок идет медленно и через некоторое время прекращается. Эти механизмы управляются на молекулярном уровне генами самонесовместимости. Самонесовместимость, широко распространена, она встречается примерно у 10000 видов цветковых растений из 78 семейств.
Гетеростилия (греч. heteros – другой, stylos — столб), или разностолбчатость, представляет собой развитие на одной особи цветков с различной высотой тычинок и столбиков. Это приводит к тому, что в основном опыляются цветки разных морфологических типов, т. е. пыльца с длинностолбчатых цветков переносится, главным образом, на короткостолбчатые цветки и наоборот – с короткостолбчатых цветков большая часть пыльцы попадает на длинностолбчатые. Опыление цветков сходного типа при этом также происходит, но очень редко. Разумеется, длина тычинок и столбиков контролируется генами, причем их локусы в хромосоме расположены очень близко и наследуются вместе. Гетеростилия встречается относительно редко (обнаружена у 56 родов из 23 семейств).
Перечисленные адаптации являются наиболее известными, но не единственными. У разных покрытосеменных существует большое количество разнообразных морфологических приспособлений, например строение цветка, его ориентация в пространстве и т. д.Способы опыления у перекрестноопыляемых цветковых растений очень разнообразны, Условно их можно разделить на две группы. В первый перенос пыльцы осуществляется животными, главным образом, насекомыми, а также некоторыми позвоночными (большинство из которых также освоили воздушную среду – птицы и летучие мыши). Во второй группе пыльца переносится абиотическими факторами – ветром и реже водой.
У растений, опыляемых животными, обычно формируются различные адаптации, способствующие привлечению опылителей. Обычно они имеют яркоокрашенные крупные цветки. Если цветки мелкие, то они, как правило,оказываются собранными в соцветия, что также зрительно увеличивает их.
Признаки насекомоопыляемых растений
1. Имеются нектарники, железы в основании пестика, которые вырабатывают сладкий нектар. На эту вкусную жижицу, да еще «приправленную» пыльцой, охотно слетаются насекомый. Как следует покопошившись в цветке, опылив его, они, все в пыльце, летят на другие цветки и тоже их опыляют.
2. Цветки обоеполые, пыльца липкая, клейкая, на оболочке пыльцы имеются выросты. Обоеполость цветков нужна, чтобы опыление насекомым происходило сразу, на этом же цветке. А клейкость пыльцы и выросты — для прикрепления к покрову насекомого.
3. Цветки зачастую крупные, их околоцветник ярко, привлекательно окрашен. Это приспособление выработано в рекламных целях — яркие цветки больше привлекают насекомых.
4. Если цветки мелкие, то они собраны в крупные соцветия — для тех же целей.
5. Цветки имеют выраженные запахи. Неизвестно, какие запахи больше нравятся насекомым, но человек различает среди них приятные сладкие запахи (липа, сирень, гиацинт, роза) и отталкивающие, — например, раффлезия Арнольда распространяет запах гнилого мяса, а аморфофаллус титанический (наибольший цветок в мире) «благоухает» протухшими яйцами и рыбой. Однако же и на таких красавцев находятся свои опылители! Но нам все-таки приятнее какой-нибудь душистый табак, чей чудесный аромат разливается в летнюю ночь.
Приспособленность цветков к опылению определенными насекомыми
Многие растения «накрывают стол» для любых насекомых, у них пыльца и нектар доступны всем, кто прилетит, как, например, у мака. Но некоторые виды опыляются только определенными насекомыми. Так, сердцевина цветка львиного зева труднодоступна — в него еще пролезть нужно! И сделать это могут только крупные увесистые насекомые: шмели и пчелы. Они своим тельцем приоткрывают цветок и проникают внутрь. А вот упомянутый уже душистый табак опыляют крупные ночные бабочки бражники, — их очень длинный и тонкий, как игла, хоботок дотягивается до нектара и пыльцы. В тропиках опылением занимаются даже летучие мыши и птицы — например, крошечные колибри.
Признаки ветроопыляемых растений
1. Деревья и кустарники цветут еще до появления первых листьев. Пока нет листвы, леса и парки голые, пыльце легче циркулировать, нестись за ветром. Так ведут себя, например, шелковица, ольха и орешник.
2. Не имеют околоцветника, а если он есть, то развит слабо (злаки).
3. Цветки мелкие, зачастую собраны в соцветия (ива, пшеница).
4. При таком виде опыления пыльца нередко расходуется впустую, улетает без толку, поэтому растение вырабатывает ее с большим избытком. Крупные и клейкие пылинки также не имеют смысла — прилипнут неподалеку, и поэтому здесь пыльца очень мелкая, сухая, летучая.
5. Одиноко стоять на равнине и ждать опыления очень грустно, поэтому ветроопыляемые растения предпочитают образовывать скопления (березовые рощи, тростниковые заросли).
1. Этот вид опыления, перенос пыльцы с женских цветков на мужские, вручную делает человек в определенных целях — выведение новых сортов, подъем урожайности.
2. Как производится опыление? С мужских цветков удаляют пыльники, подсушивают пыльцу и наносят ее на женские цветки в определенное время — причем обрабатывать вообще все цветки в теплице, например, не требуется, достаточно охватить десяток процентов. Еще один способ — аккуратно срезать мужские цветки и потрясти их пыльниками над женскими, стараясь не повреждать последние. Например, однодомную кукурузу, имеющую мужские соцветия-метелки и женские початки, можно опылить так, не ожидая милостей от природы: собрать в бумажную воронку пыльцу и рассеять ее над рыльцами пестиков.
Что ты хочешь узнать?
Ответ
У растений, опыляемых ветром, цветки диаметрально противоположны цветкам, которые опыляются насекомыми. Ветер – стихийный фактор и может нести пыльцу в разных направлениях. Чтобы использовать его, растениям необходимы совсем другие цветки, как при опылении насекомыми. При опылении ветром нет надобности тратить ценные материалы на яркую окраску цветочных покровов, на образование сладкого нектара, душистого аромата. Здесь вырабатывались другие приспособления, направленные в сторону упрощения строения цветка.
Поэтому цветки ветроопыляемых (анемофильных) растений невзрачны, не издают никакого запаха, не выделяют нектара. Околоцветник у них очень слабо развит или совсем отсутствует. Он здесь и не нужен. Напротив, выдвинутые далеко наружу пыльники свободно овеваются ветром (злаки, осоки) , который выдувает из них пыльцу и рассеивает по воздуху. Даже легкий ветерок качает сережки, метелки, тычинки.
Наши деревья и кустарники (тополь, орешник и др. ) цветут обычно весной, когда дуют сильные ветры и еще не распустилась листва, так что ветер без помехи сдувает пыльцу на цветки. Опыляемые ветром растения не растут в одиночку, а образуют большие заросли, что также увеличивает шансы опыления их цветков. Ветер рассеивает бесполезно очень много пыльцы, поэтому растения образуют ее в огромных количествах. Например, в сережке обыкновенной лещины насчитывается до миллиона пыльцевых клеток. А когда цветет сосна, то в воздухе поднимаются целые облака желтой пыльцы, которая оседает на землю в виде так называемого серного дождя. Пылинки сосны к тому же имеют специальные приспособления для летания в виде двух воздушных шаров. И вообще у всех ветроопыляемых растений пыльца мелкая, легкая, сухая. Благодаря этому ветер легко выдувает ее из пыльников. И рыльца в свою очередь хорошо приспособлены к улавливанию пыльцы. Так же как и пыльники, в период цветения они выставляются далеко наружу и имеют вид густых перьев (злаки) , длинных нитей (кукуруза, осоки) или кисточек (лещина)